veröffentlicht in “Der Vogelfreund” 2/2005
Carotinoide
sind wichtige Stoffe für den Vitamin-A-Haushalt und für die Lipochromentwicklung
Was sind Carotinoide?
Carotinoide sind als natürliche Pigmente in der Natur weit verbreitet. Wir kennen sie als gelbe bis rote Naturfarbstoffe von den Tomaten, dem Paprika, dem Mais, den Karotten und vom bunten Herbstlaub. Carotinoide färben jedoch auch die Panzer der Krebse, das Fleisch der Lachse, das Eigelb der Vögel und natürlich – was uns hier besonders interessiert – die Federn vieler Vogelarten.
Was sind Lipochrome?
Die Bezeichnung „Lipochrom“ stammt noch aus der Mitte des 20. Jahrhunderts, als der Begriff „Carotinoid“ noch nicht festgelegt war. Man bezeichnete die in Fetten löslichen Pigmente als Lipochrome (griech.: lipos = Fett; chroma = Farbe) oder zu Deutsch: „Fettfarbe“. Später wurde international der Begriff „Carotinoid“ (auch „Karotinoid“) für diese große Stoffgruppe eingeführt.
Nur in der Kanarienzucht hat sich seit mehr als 50 Jahren dieser alte Begriff bis zum heutigen Tag erhalten!
Wir Kanarienzüchter sprechen vom Lipochrom oder von der Fettfarbe, wenn wir die Grundfarbe des Gefieders unserer Kanarien meinen. So gesehen hat das „Lipochrom“ eine neue Begrifflichkeit erhalten und kann dann – aber bitte nur dann – für die Gefiederfärbung unserer Vögel gelten.
Wir wissen jedoch, dass die „Lipochrome“ eigentlich Carotinoide sind, oder noch exakter Xanthophylle!
Etwas Chemie muss sein!
Die Carotinoidmoleküle bestehen vor allem aus Kohlenstoff (C) und Wasserstoff (H) und sind demnach Kohlenwasserstoffe. Diese beiden Elemente werden zu große Molekülen zusammengesetzt. Meist bestehen die Moleküle der Carotinoide aus 40 Kohlenstoffatomen und 56 Wasserstoffatomen (C40H56).
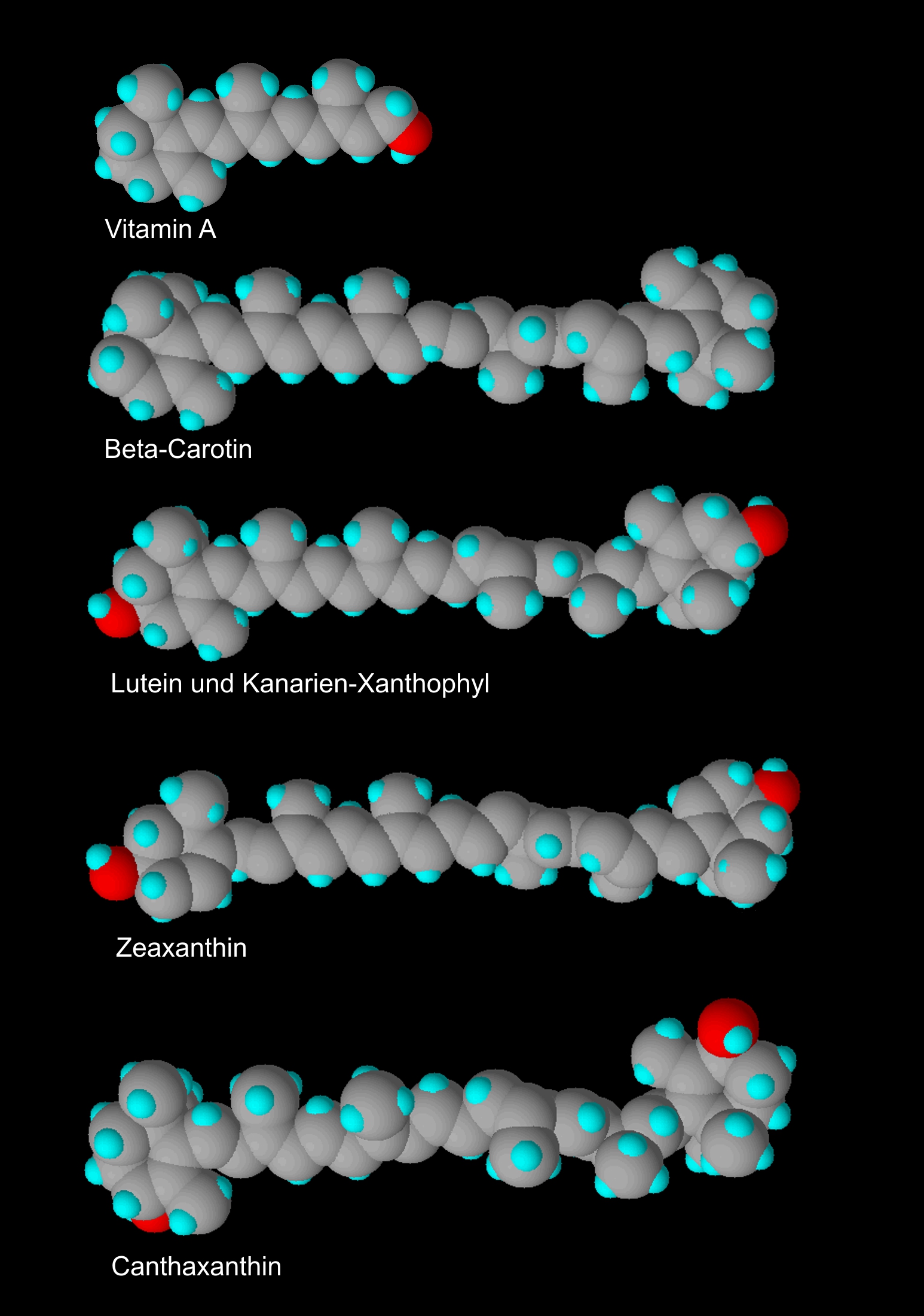
Die Carotinoide gehören zu den Polyenen, also Stoffen, die in ihrer chemischen Struktur viele Doppelbindungen aufweisen („poly“ = viel, „-en“ bezeichnet eine Doppelbindung). Die Größe des Molekülgrundgerüstes und die Anzahl der Doppelbindungen bestimmen die Lichtabsorption. D. h. je mehr Doppelbindungen vorhanden sind, umso gelber bzw. röter erscheint uns die Farbe. Dabei bleibt die Anzahl der Atome relativ konstant. Es ändert sich nur die Atomanordnung innerhalb des Moleküls. Solche Moleküle, die sich nur in der Atomanordnung unterscheiden, nennt man Isomere.
Die beiden Enden dieser langen, unverzweigten Molekülkette besitzen Endgruppen, die zu Ringen geformt sein können. An diesen Endgruppen können noch andere Elemente angefügt sein. Die Molekülenden haben – je nach Carotinoid – einen bestimmten Aufbau und verfügen über weitere Doppelbindungen an
unterschiedlichen Stellen. Zur besseren Unterscheidung werden diese Endgruppen nach Buchstaben des griechischen Alphabets bezeichnet (Alpha, Beta, Gamma…). Der Aufbau dieser Molekülringe hat wesentlichen Einfluss auf die weitere Synthesekette der Carotinoide.
Besitzt das Molekül zwei gleichartige Ringe, wird es als symmetrisch bezeichnet (z. B. ß,ß). Sind die Ringe unterschiedlich (z. B. ß,e) sprechen wir von einem asymmetrischen Aufbau.
Die Carotinoide werden in die sauerstofflosen Carotine und die sauerstoffhaltigen Xanthophylle unterschieden.
Provitamin A
In Pflanzen liegen Carotinoide als ein – je nach Pflanzenart unterschiedlich – Gemisch aus verschiedenen Carotinen und Xanthophyllen vor. Pflanzen können Carotinoide synthetisieren, aber kein Vitamin A erzeugen!
Pflanzenfressende Tiere – also auch unsere Kanarienvögel – können mit der natürlichen Nahrung kein oder nur wenig Vitamin A aufnehmen. Der Tierkörper muss das Vitamin selbst in seinem Körper herstellen. Dazu benötigt er bestimmte Carotinoide, die Provitamine A, die in den Pflanzenteilen enthalten sind.
Die wichtigsten Provitamine A sind für uns Alpha-Carotin, Beta-Carotin und Gamma-Carotin. Sie besitzen mindestens eine ß-Endgruppe ohne Sauerstoffatom.
Vitamin A
Die Provitamine A lassen sich in der Dünndarmschleimhaut in zwei gleichlange Teile zerlegen. Zum Beispiel das Beta-Carotin (ß,ß-C40H56) in zwei ß-C20H28)-Teile. Diese ß-C20H28)-Teile können durch Anlagerung von Sauerstoff und – in einem zweiten Schritt – Wasserstoff zu ß-C20H30O (= Vitamin A1 = Retinol) umgewandelt werden. Das Vitamin A1 wird im weiteren Syntheseverlauf verestert, und kann dann als Speicherform in der Leber deponiert werden.
Das Alpha- und das Gamma-Carotin besitzen nur eine ß-Endgruppe und können deshalb nur ein Molekül Vitamin A1 bilden.
Die Fähigkeit, aus Carotinen (Provitamine A) Vitamin A bilden zu können, muss durch ein oder mehrere Enzyme – und damit durch ein oder mehrere Gene – gesteuert werden.
Unsere Vögel benötigten täglich eine bestimmte Menge Vitamin A, das als Provitamin A in der Nahrung enthalten sein muss. Durch die Speicherung von Vitamin A in der Leber können Schwankungen des Provitamin-A-Gehaltes der Nahrung ausgeglichen werden.
Vitamin A ist für Vögel lebensnotwendig, denn es regelt viele Körperfunktionen und Lebensvorgänge. Bei Vitamin-A-Mangel können folgende Störungen auftreten:
- Rückgang der Legetätigkeit,
- vermehrt unbefruchtete Eier und schlechte Schlupfergebnisse,
- Wachstumsstörungen bei jungen Vögeln,
- Augenentzündungen und Erblindung,
- Beeinträchtigung der Federausbildung,
- allgemeine Anfälligkeit gegen Krankheitserreger.
Ein lang anhaltender Vitamin-A-Mangel hat letale (tödliche) Wirkung. Dabei ist es gleichgültig, ob in der Nahrung die Provitamine A fehlen, oder ob der Vogel, durch einen genetischen Defekt, nicht in der Lage ist Vitamin A zu entwickeln.
Auch ein Zuviel an Vitamin A kann Störungen hervorrufen:
- Wachstumsstörungen,
- Auswirkungen auf das Knochensystem,
- schwere Schädigung der Federbildung
Ein Überangebot an Vitamin A kann nur der Züchter herbeiführen, indem er zu viel reines Vitamin A verfüttert.
Ein Zuviel von Provitaminen A in der Nahrung hat jedoch keine Nachteile, da der Körper nur eine bestimmte Menge Provitamin A in Vitamin A umsetzen kann. Der weitere Überschuss wird ausgeschieden.
Lutein – der universelle Stoff zur Bildung der Lipochrome
Lutein ist ein asymmetrisch aufgebautes Xanthophyll von kräftig gelber Farbe. Lutein hat die Bruttoformel ß,e-C40H56O2.
Es entspricht somit im Aufbau dem Alpha-Carotin, denn es besitzt die Endgruppen ß und e und die, bereits von anderen Carotinoiden bekannten, 40 Kohlenstoffatome (C40) und 56 Wasserstoffatome (H56). Die zwei Sauerstoffatome (O2) sind als Hydroxyl mit einer Einfachbindung an beiden Endgruppen angelagert (siehe Strukturformel).
Lutein kommt sehr häufig in Pflanzenteilen vor, aber auch im Gefieder mancher Vögel und im Eidotter.
Völker [4] hat bereits 1962 Fütterungsversuche mit gelb- und rotgrundigen Kanarien durchgeführt. Er konnte nachweisen, dass die gelbe und – mit Einschränkung – rote Farbe des Gefieders (das „Lipochrom“) nur durch Lutein in der Nahrung entwickelt werden kann.
Umwandlung von Lutein in gelbes Lipochrom
Völker´s Fütterungsversuche ergaben, dass nur durch Verfütterung von gelbem Lutein ein körpereigener gelber Gefiederfarbstoff entsteht. Dass mit der Nahrung aufgenommene Lutein wird demnach im Vogelkörper zu einem speziellen Farbstoff, dem Kanarienxanthophyll, umgewandelt.
Da dieser spezielle Farbstoff zuerst bei Kanarien beschrieben wurde, nannte man ihn „Kanarienxanthophyll“. Wie der Name bereits aussagt, handelt es sich um ein Xanthophyll, also um ein sauerstoffhaltiges Carotinoid.
Heute wird das Kanarienxanthophyll meist nur Xanthophyll genannt. Der Molekülaufbau entspricht dem Lutein, hat also ebenfalls die Bruttoformel ß,e-C40H56O2. Das Kanarienxanthophyll ist somit ein Isomer des Luteins. Lutein und Kanarienxanthophyll unterscheiden sich nur an Hand der Spektralabsorbtion. Die Wellenlänge des zurückgeworfenen Lichtes wird beim Xanthophyll mit 442 nm und beim Lutein mit 445,5 nm angegeben. Aufgrund der großen Ähnlichkeit von Lutein und Xanthophyll werden beide Begriffe gern als Synonyme verwendet.
Die Fütterungsversuche Völker´s ergaben weiterhin, dass auch das Xanthophyll Zeaxanthin zur Gelbfärbung der Kanarien geeignet ist (bei ausschließlicher Zeaxanthinfütterung im Versuch). Das Zeaxanthin kommt in der Nahrung nur in sehr geringer Menge vor. Die vom Vogel aufgenommene Menge reicht deshalb allein nicht aus, um eine Gelbfärbung zu erreichen.
Zeaxanthin besitzt die Struktur ß,ß-C40H56O2 und wird – wie das Lutein – vom gelbgrundigen Kanarienvogel ebenfalls zum körpereigenen Kanarienxanthophyll umgewandelt.
Ich konnte leider bisher nicht herausfinden, auf welche Weise bei Tieren oder Vögeln das Lutein und das Zeaxanthin in körpereigene Lipochrome umgewandelt werden. Möglicherweise werden (analog der Vitamin-A-Synthese) alle aufgenommenen Carotinoide in zwei Teile gespalten. Anschließend können die Teilstücke der Provitamine A zu Vitamin A, und die Teilstücke bestimmter Xanthophylle zu Lipochromen neu zusammengesetzt werden. Sicher ist, dass das Lutein und das Zeaxanthin in einer Synthese zu einem neuen Molekül umgewandelt werden, dem Kanarienxanthophyll.
Die Fähigkeit der gelbgrundigen Kanarien, aus Lutein und Zeaxanthin Kanarienxanthophyll bilden zu können, muss durch ein oder mehrere Enzyme – und damit durch ein oder mehrere Gene – gesteuert werden.
Häufig wird darauf verwiesen, dass auch Beta-Carotin für eine Gelbfärbung verantwortlich ist. Völker untersuchte auch das und stellte fest, dass Beta-Carotin keinerlei Gelbfärbung herbeiführt. Beta-Carotin kann also nur zu Vitamin A umgewandelt werden. Allerdings hat dieses Vitamin Einfluss auf die Lebensfunktion und Auswirkungen auf die Vitalität und somit auch auf ein optimal ausgebildetes Gefieder.
Umwandlung von Lutein in rotes Lipochrom
Das rote Lipochrom des Kanarienvogels ist durch den Kapuzenzeisig in die damals nur gelben (und weißen) Kanarien übertragen worden.
Das rote Gefieder des Kapuzenzeisigs wird durch Canthaxanthin hervorgerufen. Canthaxanthin ist ein symmetrisches Xanthophyll mit ß,ß-Molekülendgruppen. An diesen beiden Endgruppen ist jeweils ein Sauerstoffatom mit einer Doppelbindung angekoppelt (siehe Strukturformel).
Völker stellte durch seine Fütterungsversuche fest, dass das rote Lipochrom durch Umwandlung des gelben Luteins erzeugt wird (auf eine Ausnahme komme ich später zurück).
Der gelbe Ausgangsstoff Lutein wird demnach zu zwei farblich sehr unterschiedlichen Lipochromen umgewandelt. Aus Lutein wird beim gelbgrundigen Kanarie das Kanarienxanthophyll gebildet und aus dem gleichen Lutein beim rotgrundigen Kanarie Canthaxanthin. Betrachten wir uns die Strukturformeln, so stellen wir fest, dass nur die Doppelbindungen mit Sauerstoff entscheiden, ob aus Lutein gelbes oder rotes Lipochrom entsteht!
Aus Völker´s Untersuchungen wissen wir, dass die rote Gefiederfarbe des Kapuzenzeisigs auchGelbanteile aufweist. Er konnte diesen geringen Gelbanteil als Kanarienxanthophyll bestimmen. Im Sichtbild wird der geringe Gelbanteil vom optisch stärkeren Rotanteil überdeckt. Es ist also anzunehmen, dass der Kapuzenzeisig beide Xanthophylle (gelbe und rote) gleichzeitig aber in unterschiedlicher Menge synthetisiert.
Die Fähigkeit des Kapuzenzeisigs und der rotgrundigen Kanarien, aus Lutein Canthaxanthin und Kanarienxanthophyll bilden zu können, muss durch ein oder mehrere Enzyme – und damit durch ein oder mehrere Gene – gesteuert werden.
Umwandlung von Beta-Carotin in rotes Lipochrom
Völker stellte weiterhin fest, dass durch alleinige Verfütterung von reinem Beta-Carotin rotgrundige Kanarien tatsächlich rotes Lipochrom ausbilden können. Das Rot war jedoch in seiner Farbwirkung wesentlich schwächer.
Biochemisch betrachtet, muss „nur“ dem ß-C20H28-Teil des Beta-Carotins ein Sauerstoffatom mit einer Doppelbindung angefügt werden. Offensichtlich geschieht das nicht mit jedem Beta-Carotin-Molekül, da sonst kein Unterschied in der Farbtiefe festzustellen wäre.
Die Fähigkeit des Kapuzenzeisigs und der rotgrundigen Kanarien, aus ß-Carotin rotes Lipochrom bilden zu können, muss durch ein oder mehrere Enzyme – und damit durch ein oder mehrere Gene – gesteuert werden.
Die Wirkung roter Xanthophylle auf die Lipochrome gelbgrundiger Kanarien
Wir füttern manche gelbgrundigen Positurkanarien mit roten Farbstoffen, um im Gefieder orangefarbenes Lipochrom zu erzielen.
Früher wurde der Farbstoff des Paprikas, das Capsanthin (ein Xanthophyll mit einer Sauerstoff-Doppelbindung), dazu verwendet. Heute wird vielfach das synthetisch hergestellte Canthaxanthin dafür eingesetzt.
Gelbgrundige Kanarien sind genetisch nicht in der Lage, rotes Lipochrom selbst zu erzeugen. Sie können aber offensichtlich die „fertigen“ roten Xanthophylle ohne weitere Umwandlung in der Feder einlagern. Da diese gelbgrundigen Vögel jedoch noch die volle Gelbsynthese durchführen, werden die roten Farbstoffe nur zusätzlich in der Feder eingelagert. Der Anteil roter Farbstoffe ist nicht so hoch, dass eine rote Gefiederfarbe entsteht. Sie bleibt lediglich orange.
Die möglichen genetischen Voraussetzungen der Lipochrombildung
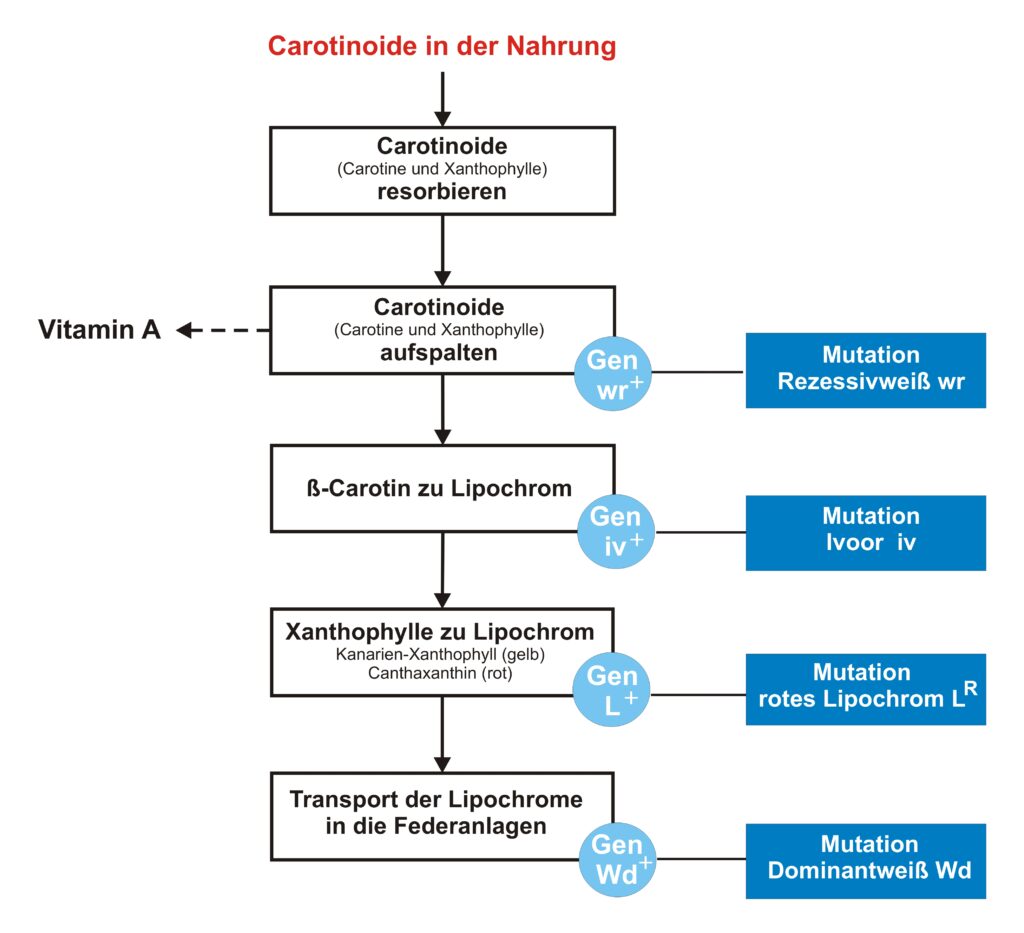
Mutation Rezessivweiß
Der Vogel muss Carotinoide mit der Nahrung aufnehmen, um alle weiteren Stoffwechselvorgänge ausführen zu können. Werden ihm (in Fütterungsversuchen) alle Carotinoide entzogen, kann er weder Lipochrome noch Vitamin A bilden. Der Vitamin-A-Mangel führt unweigerlich zum Tod.
Die aufgenommenen Carotinoide werden von der Dünndarmschleimhaut aufgesogen (resorbiert). Dafür wird bisher in der Kanarienliteratur ein Karotinaufnahmefaktor oder auch Karotinresorbtionsfaktor „K“ (also ein Gen) verantwortlich gemacht. Das Fehlen dieses Faktors würde den Rezessivweißen hervorrufen. Das rein physikalische Aufsaugen (lat. = resorbieren) von Nahrungsbestandteilen kann m.E. nicht an ein Gen gebunden sein. Ein Faktor „K“ hat also an dieser Stelle nichts zu suchen. Der Begriff ist unglücklich gewählt.
Die resorbierten Carotinoid-Moleküle werden in der Dünndarmschleimhaut in zwei Teile gespalten (siehe obige Ausführungen). Nur diese Molekülteile stehen für weitere Synthesen zur Verfügung.
Die Aufspaltung der Carotinoide wird durch ein oder mehrere Enzyme hervorgerufen. Diese Enzyme wiederum werden durch ein bzw. mehrere Gene gesteuert (Ein-Gen-ein-Enzym-Hypothese). Tritt hier eine genetische Störung (= Mutation) auf, so sind alle weiteren Syntheseschritte gehemmt oder können überhaupt nicht ablaufen. Ein Vogel mit dieser Störung kann weder Vitamin A noch Lipochrome bilden und ist aufgrund des Vitamin-A-Mangels nicht lebensfähig. Diese Mutation kennen wir beim Kanarienvogel als Rezessivweiß (Englischweiß).
Da die bisherige Faktorenbeschreibung nicht den Tatsachen entspricht, kann auch das Symbol „K“ nicht verwendet werden. Ich bezeichne diese Mutation als „wr“ (weiß oder withe und rezessiv). Daraus ergeben sich folgende Phänotypen:

Mutation Ivoor
Aus Völkers Fütterungsversuchen wissen wir, das rotgrundige Kanarien Beta-Carotin zu rotem Lipochrom (Canthaxanthin?) umwandeln können. Zitat: „Die Rotvögel reagieren auf Carotinfütterung mit einer zarten Rosafärbung ihrer Federn, die sich durch ihren ausgeprägt roten Farbton deutlich von jenem der Originalfedern unterscheidet. Die Farbstufe liegt etwa bei OF 5 ea – 5 ¼ ea (= ,besonders blaß’).“ Da Völker keine Angaben darüber machte, ob sich diese Rotfärbung nur auf bestimmte Gefiederzonen beschränkt (Mosaik), muss diese zarte Rosafärbung den ganzen Körper umfasst haben.
Eine zarte Rosatönung kennen wir von den Rot-Ivoorvögeln, die früher als „Lipochrom-Pastell“ bezeichnet wurden. Henniger gibt dazu an: „Bei ihr (dieser Farbspielart N.S.) ist die jeweilige Fettfärbung ganz gleichmäßig über den ganzen Körper abgeschwächt, … In meiner ,Ostwaldschen Farbnormen-Meßtafel’ entspricht die dritte Gruppe (besonders blaß) durchaus den aufgehellten Färbungen der Spielart ,Pastell’“.
Die Möglichkeit, aus Beta-Carotin rotes Lipochrom zu synthetisieren, muss an einem Erbfaktor gekoppelt sein. Da diese Möglichkeit nur an den damaligen Rotvögeln festgestellt wurde, muss diese Eigenschaft vom Kapuzenzeisig stammen und auf die Kanarien übertragen worden sein.
Da es auch gelbgrundige Ivoorvögel gibt, wäre es sehr interessant, ob diese auf reine Carotin-Fütterung mit einer pastellgelben Lipochromfärbung reagieren, wie damals die rotgrundigen Vögel.
Wir wissen, dass „Ivoor“ geschlechtsgebunden rezessiv vererbt. Die Erbformel lautet „iv+“.
Mutation rotes Lipochrom
Die gelbgrundigen Kanarien verarbeiten die Xanthophyllteilstücke zu dem körpereigenen Kanarienxanthophyll. Dafür sind ebenfalls ein oder mehrere Gene und Enzyme verantwortlich. Diese Umwandlung zu Kanarienxanthophyll ist bisher noch keiner Mutation unterworfen worden. Deshalb wird als Erbformel „L+“ (Lipochrom gelb) gewählt.
Die frühere Bezeichnung „g“ für nicht vorhandenes Gelb ist nicht haltbar. Nicht vorhandenes Gelb (= kein Lipochrom) wird durch wr oder Wd hervorgerufen.
Die rotgrundigen Kanarien verarbeiten die Xanthophyllteilstücke zu rotem Canthaxanthin. Das Lutein wird hier nicht zu Gelb, sondern zu Rot synthetisiert. Das bedeutet, dass lediglich das dafür verantwortliche Enzym – und damit sein Gen (= Mutation) – verändert ist. Somit ist beim Kapuzenzeisig das Enzym für die Gelbausfärbung mutiert (aus „L+“ wird „LR“ – Lipochrom rot) und kann nunmehr rotes Lipochrom erzeugen. Der Genort für die Lipochromsynthese ist also beim Kanarienvogel und beim Kapuzenzeisig gleich.
Die frühere Bezeichnung „r“ für nicht vorhandenes Rot ist nicht haltbar. Nicht vorhandenes Rot (= kein Lipochrom) wird durch wr oder Wd hervorgerufen.
Eine Kreuzung zwischen gelben und roten Kanarien ergeben orangefarbene Nachkommen. Diese haben aus Lutein sowohl Kanarienxanthophyll als auch Canthaxanthin gebildet und gemeinsam in die Feder eingelagert.
Mit dieser Tatsache ist es sehr verwunderlich, dass aus einer Selbstung orangener Kanarien u.a. „vollweiße“ Kanarien entstehen sollen, d. h. Kanarien, die keinerlei Lipochrome – weder Kanarienxanthophyll noch Canthaxanthin – bilden können. Nach unserem Ausflug in die Biochemie wissen wir jetzt, das das nicht möglich sein kann. Auf die Rot-Gelb-Vererbung gehe ich in einem anderen Artikel ein.
Mutation Dominantweiß
Wir kennen in der Lipochromreihe noch eine weitere Form – die Dominantweißen (genauer die einfaktorigen Dominantweißen). Diese zeigen, gemäß unserem Standard, noch Reste von gelben bis roten Lipochrom in den Schwungfedern. Es treten auch hin und wieder Exemplare auf, die an anderen Stellen des Gefieders Lipochromflecken zeigen oder mit einem leichten Hauch an Lipochromfarbe überzogen sind. Das beweist, dass auch einfaktorige dominantweiße Kanarien genetisch in der Lage sind, Lipochrom zu erzeugen; und zwar das gesamte Spektrum von gelb bis rot sowie gelbivoor bis rotivoor.
Es wird vermutet, dass das Lipochrom während der Federentwicklung nicht in alle Federteile gelangen kann. Nur in die zuerst gebildeten Federn gelangt noch Lipochrom. Beziehen wir die allgemeine Erkenntnis ein, dass Erbfaktoren mitunter eine bestimmte Zeit brauchen, bis sie voll wirksam sind, dann lässt sich auch erklären, warum die bewussten Federpartien Lipochromeinlagerungen haben, später wachsende Federn jedoch nicht.
Dem Dominantweißen hat man den Faktor „F“ (Fettfarbentwicklungsfaktor) zugeordnet. Wir wissen jedoch, dass sie Lipochrome durchaus entwickelt können. Deshalb verwende ich, in Analogie zum Rezessivweißen die Formel „Wd+“ (weiß oder withe und dominant).

Bleibt noch eine offene Frage zu klären: Warum sind doppelfaktorige Dominantweiße nicht lebensfähig (letal)? Allein das vollständige Fehlen der Lipochrome kann m. E. nicht zum Tode führen.
Quellen undLiteratur
Henniger, J.: Farbenkanarien. Ein Lehrbuch für Farbenkanarienzüchter insbesondere über Farbenvererbung. Maximiliansau 1962.
Lexikon der Biochemie. Elsevier GmbH München; Spektrum Akademischer Verlag Heidelberg-Berlin 1999.
Völker, O.: Die Abhängigkeit der Lipochrombildung bei Vögeln von pflanzlichen Carotinoiden. J.Orn. 1934.
[4] Völker, O.: Experimentelle Untersuchungen zur Frage der Entstehung roter Lipochrome in Vogelfedern; Farbfütterungsversuche am Roten Kanarienvogel. J.Orn. 1962.