Veröffentlicht im „Der Vogelfreund“ 6/2005
Die Vererbung gelber und roter Lipochrome
Historie
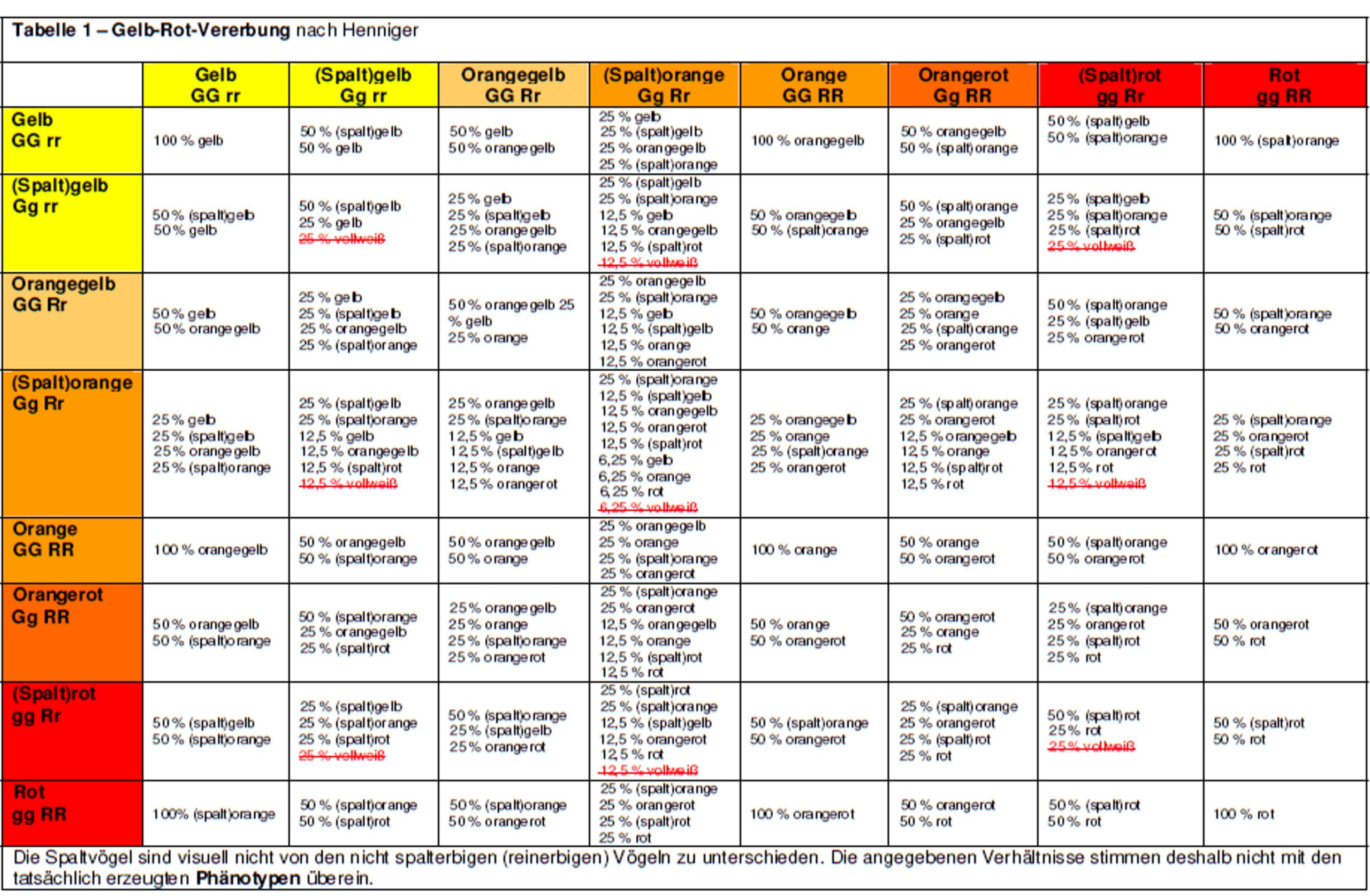
Der Altvater der Farbenkanarienzucht, Julius Henniger [3], hat 1962 ein System der Erbfaktoren erstellt, mit dem die folgenden Züchtergenerationen ein wichtiges Rüstzeug für die Kanarienzucht in die Hände bekamen.
Henniger ging davon aus, dass die Gelbvögel ein Gen für die Gelbausfärbung und die Kapuzenzeisige ein anderes Gen für die Rotausfärbung besitzen. Deshalb hat er für den gelben Phänotyp des Kanarienvogels den Faktor „G“ und für den roten Phänotyp den Faktor „R“ festgelegt. Da Gelbvögel kein Rot besitzen, musste das mit einem „r“ bezeichnet werden. Das fehlende Gelb beim Rotvogel bezeichnete er mit „g“. Der reinerbige Gelbvogel hatte nach Henniger die Erbformel „GG rr“ und der reinerbige Rotvogel die Formel „gg RR“.
Kreuzt man gelbe mit roten Kanarien, so sind alle Nachkommen einheitlich (uniform) orangefarben (Henniger – ein Feind von Fremdwörtern in der deutschen Sprache – bezeichnete diese Farbe als „Kreß“, benannt nach der Farbe der Kapuzinerkresse). Nach dem 1. Mendelschen Gesetz (Uniformitätsgesetz) deutete dieses Ergebnis auf einen intermediären (mittelnden) Erbgang hin, denn alle Nachkommen waren gleich (uniform) in der Farbe. Sie mussten demnach mischerbig sein. Henniger nannte diese orangefarbenen Nachkommen „Spaltkreß“ (= Spaltorange). Auch die errechnete Erbformel dieser orangefarbenen Nachkommen mit der Erbformel „Gg Rr“ schienen die Richtigkeit aller bisherigen Annahme zu bestätigen.
Verpaart man die orangefarbenen Nachkommen untereinander, sind deren Nachkommen sehr vielfältig in der Lipochromfärbung, denn es wirkt das 2. Mendelsche Gesetz (Spaltungsgesetz). Von gelb über orangegelb, orange, orangerot bis rot sind alle möglichen Farbnuancen der Lipochrome vorhanden. Alle Ergebnisse der Gelb-Rot-Verpaarungen, die sich aus dem Henniger-System ergeben, habe ich in Tabelle 1 zusammengefasst.
Biochemische Vorgänge der Lipochromentwicklung
- gelbgrundige Kanarien bilden aus dem Xanthophyll „Lutein“ das körpereigene „Kanarien-Xanthophyll“;
- Kapuzenzeisige und rotgrundige Kanarien bilden aus dem Xanthophyll „Lutein“ das körpereigene Xanthophyll „Canthaxanthin“;
- Kapuzenzeisige lagern gelbes „Kanarien-Xanthophyll“ und rotes „Canthaxanthin“ gemeinsam in der Feder ab. Die Menge des roten Canthaxanthin ist deutlich größer als die Menge des gelben Kanarien-Xanthophyll. Somit überdeckt das Canthaxanthin phänotypisch das Kanarien-Xanthophyll.[5] [6]
Neue Überlegungen
Die rote Lipochromfarbe der Cardueliden ist, entwicklungsgeschichtlich betrachtet, eine noch recht junge Gefiederfarbe. Sie muss sich aus Stammformen entwickelt haben, die ursprünglich nur gelbes Lipochrom bilden konnten. Das bedeutet, dass
- für die Synthese des gelben Gefiederfarbstoffes „Kanarien-Xanthophyll“ aus Lutein ein entsprechendes Gen (L+) – und damit ein entsprechendes Enzym – notwendig ist.
- für die Synthese des roten Gefiederfarbstoffes „Canthaxanthin“ aus Lutein ein verändertes Enzym – und damit ein verändertes Gen (= Mutation) – vorhanden sein muss.
Nach diesen Überlegungen ist beim Kapuzenzeisig das Enzym für die Gelbausfärbung mutiert (aus L+ wird Lr) und kann nunmehr rotes Lipochrom erzeugen. Der Genort für die Lipochromsynthese muss also beim Kanarienvogel und beim Kapuzenzeisig gleich sein.
Die Farbskala von Gelb zu Rot stellt sich wie folgt dar:
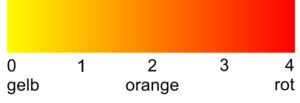
Innerhalb dieser Skala bewegen sich die Phänotypen der Kreuzungsnachkommen gelb- und rothaltiger Eltern, denn die Nachkommen werden sowohl gelbe als auch rote Lipochrome im bestimmten Verhältnis in der Feder ablagern. Ein Wegfall der Lipochrome ist nicht möglich, es kann sich nur das Verhältnis zwischen roten und gelben Lipochromen ändern. Ein (voll)weißer Vogel kann auf diese Art nicht entstehen.
Neue Theorie
Völker vermutete bereits 1962, dass die unterschiedlichen Lipochromnuancen auf einen Dosiseffekt der Enzyme zurückzuführen sind.[6] Verfolgen wir diese Vermutung mit der heutigen Kenntnis der Genetik.
In der genetischen Literatur (Brandsch, H.) wird der Dosiseffekt wie folgt beschrieben:
„Die Merkmalsausbildung zeigt quantitative Abstufungen in Abhängigkeit von der Anzahl wirksamer und unwirksamer bzw. weniger wirksamer Allele, deren Einzeleffekte sich vollständig summieren (= summatives Verhalten). Die wirkungsgleichen Gene zeigen jeweils ein intermediäres Verhalten. Unterschiedliche quantitative Effekte der betreffenden Gene führen zu entsprechend mehr Phänotypklassen der F2.“ [1]
Stellen wir uns vor, dass das Enzym mit seinem Gen L+ mehrmals in die Umbauvorgänge zum körpereigenen Kanarien-Xanthophyll eingreift. Dann würden die Teilprozesse als L1+, L2+ …. bezeichnet werden können.
Beschränken wir uns vorerst auf zwei Teilprozesse und geben jedem die Wertigkeit von 0 (= gelb).
Die Erbformel für den gelben Kanarie lautet dann „L1+ L1+ L2+ L2+“ mit einer Wertigkeit von 0 + 0 + 0 + 0 = 0 (gelb
in der obigen Farbskala).
Stellen wir uns weiterhin vor, das das mutierte Gen (L+ wird Lr ) mehrmals die Umbauvorgänge zum Canthaxanthin beeinflusst. Auch hier benennen wir Teilschritte mit L1r bzw. L2r, geben ihnen eine höhere Wertigkeit als bei gelb, z. B. 1.
Die Erbformel für den Kapuzenzeisig (roter Kanarie) lautet dann „L1r L1r L2r L2r “ mit einer Wertigkeit von 1 + 1 + 1 + 1 = 4 (rot in der obigen Farbskala).
Elterngeneration: rot L1r L1r L2r L2r x gelb L1+ L1+ L2+ L2+


Die Ergebnisse lauten zusammengefasst: 1 x gelb; 4 x orangegelb; 6 x orange; 4 x orangerot; 1 x rot.
Damit bestätigt sich die gleichsinnige Wirkung duplizierter Gene, deren charakteristisches Merkmal das Verhältnis von 1:4:6:4:1 ist!![2] [4]
Ein vollweißer Kanarie und „Spalt“-Vögel können mit dieser Erbformel nicht auftreten und bestätigen somit die praktischen Zuchtergebnisse. Die erzielten Zwischenformen entsprechen eher den praktischen Erfahrungen.
Besonders deutlich wird dies, wenn Zwischenformen miteinander verpaart werden.
In der Praxis treten weitere – hier nicht dargestellte – Farbnuancen auf. Sie lassen sich durch Hinzufügen eines 3. oder 4. Teilschrittes (L3+, L4+ bzw. L3r, L4r) durchaus darstellen. Das exakte Ansprechen jeder Farbstufe wird so aber immer schwieriger. Außerdem kann die Nuancierung auch durch die unterschiedliche Aufnahme und Verwertung des Futters hervorgerufen werden.
Beschränken wir uns auf die drei Zwischenformen orangegelb, orange und orangerot, kann das Schema wesentlich vereinfacht werden. In Tabelle 4 sind die Zusammenhänge in einfacher Form dargestellt und dürften für die züchterische Praxis ausreichen.
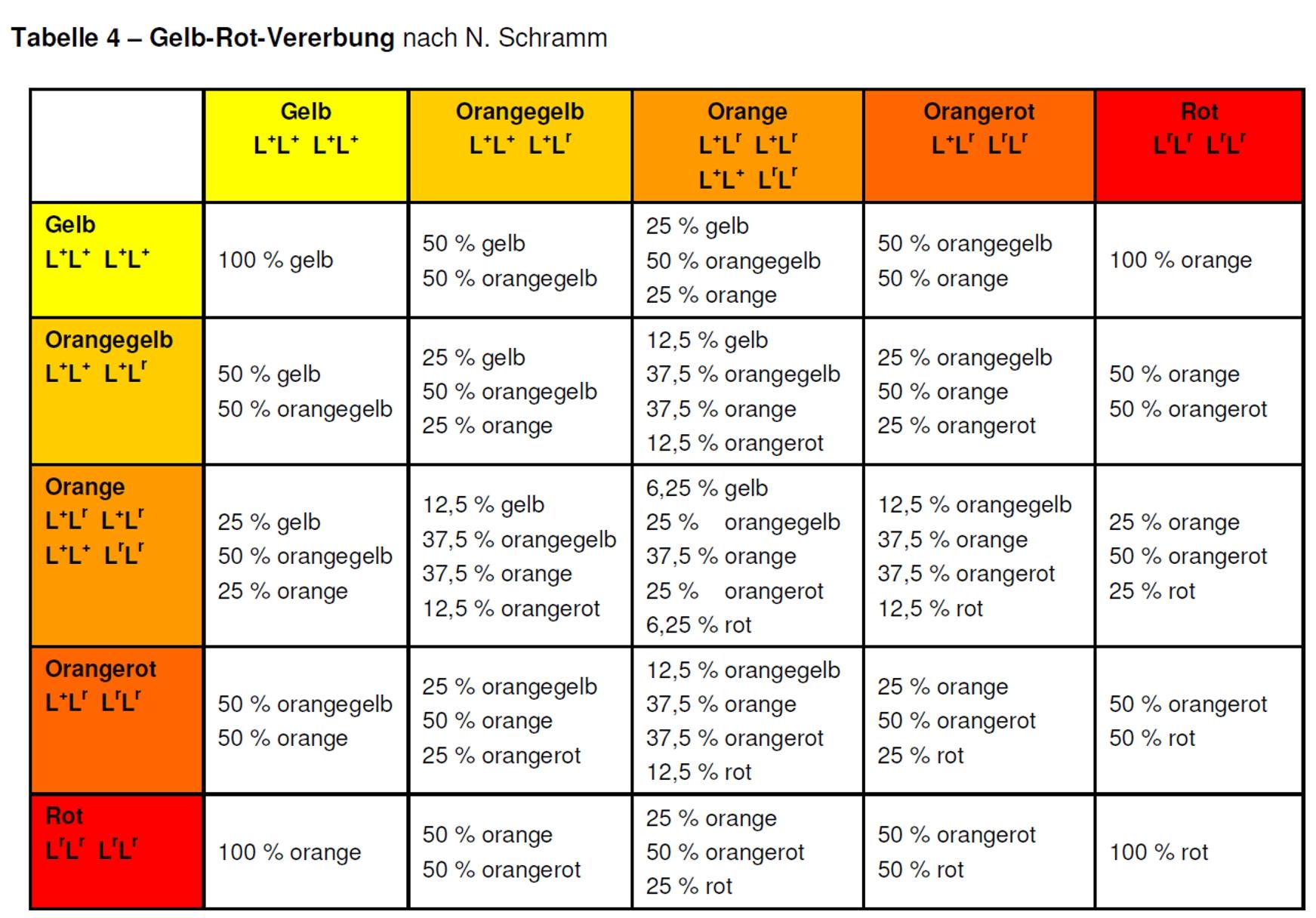
Ich hoffe, dass meine Ausführungen verständlich genug sind und ich den interessierten Lesern ein kleines Werkzeug für die praktische Zucht in die Hand geben konnte. Vor allem den Züchtern, die sich das Ziel gestellt haben, rote Gesangskanarien zu züchten, werden mit dieser Tabelle arbeiten müssen. Ich wünsche ihnen viel Erfolg auf diesem schwierigen Weg.
Quellen und Literatur
[1] Brandsch, H.: Genetische Grundlagen der Tierzüchtung. VEB Gustav Fischer Verlag, Jena 1983.
[2] Hagemann, R.: Allgemeine Genetik. VEB G. Fischer Verlag, Jena 1984.
[3] Henniger, J.: Farbenkanarien. Ein Lehrbuch für Farbenkanarienzüchter insbesondere über Farbenvererbung. Maximiliansau 1962.
[4] Seyffert, W.: Lehrbuch der Genetik. Spektrum Akademischer Verlag Heidelberg-Berlin 2003.
[5] Völker, O.: Die Abhängigkeit der Lipochrombildung bei Vögeln von pflanzlichen Carotinoiden. J.Orn. 1934.
[6] Völker, O.: Experimentelle Untersuchungen zur Frage der Entstehung roter Lipochrome in Vogelfedern; Farbfütterungsversuche am Roten Kanarienvogel. J.Orn. 1962.