
veröffentlicht auf der DKB-Homepage
Einleitung
Eine zielgerichtete Vogelzucht setzt bei jedem Züchter genetische Kenntnisse voraus. Diese Ausarbeitung soll grundlegende Kenntnisse zur Vererbung nahebringen und versteht sich nicht als wissenschaftlicher Vortrag.
Die ersten Grundgedanken zur Vererbung erhielten wir vom Augustinerpater Johann Gregor Mendel, der bereits Mitte des 19. Jahrhunderts Kreuzungsexperimente mit Pflanzen vornahm und damit ein heute noch gültiges Konzept bezüglich der genetischen Informationseinheiten, er nannte sie „Faktoren“, erstellte.
Dr. Hans Duncker unternahm in den 1930er Jahren Paarungsversuche mit Wellensittichen und Kanarienvögeln, um zu Erkenntnissen der Vererbungsweise der Gefiederfarben zu gelangen.
Das wohl bedeutendste Werk bezüglich der Farbenkanariengenetik stellt das 1962 erschienene Lehrbuch für Farbenkanarien- und Mischlingszüchter „Farbenkanarien“ von Julius Henniger dar, das zu seiner Zeit wohl als einziges Komplettwerk dieser Art bezeichnet werden konnte. Alle bis dahin bekannten Erbfaktoren handelte Henniger in diesem Werk ab.
Jedoch sind seitdem weitere 11 Mutationen (Satinet, Eumo, Onyx, Phaeo, Kobalt, Topas, Jaspe, Superoxydation, optischer Blaufaktor, Urucum und Perla) aufgetreten. Da sich diese neuen Mutationen nicht ohne weiteres in Hennigers Buchstaben-System einbauen lassen, ist eine Neuordnung des alten Systems zwingend notwendig.
Die Farbmerkmale und ihre Vererbung
Die Vielzahl an Farbmutationen bei den von uns gehaltenen und gezüchteten Vögeln steigt von Jahr zu Jahr. Um gewünschte Farbmerkmale im Stamm zu festigen und um unerwünschte Farbmerkmale zu vermeiden, ist ein Basiswissen der Genetik für alle Vogelzüchter unabdingbar.
Dieses Basiswissen gilt für alle Vogelarten und ist allgemeingültig! Beispielgebend für alle Vögel wird hier auf die Farben der Kanarienvögel eingegangen. Anhand der weiter unten aufgeführten Tabellen der Erbformeln für Kanarien und Sittiche kann die Vererbung aller bisher bekannten Gefiederfarben berechnet werden.
An dieser Stelle soll nicht auf Einzelheiten der Zellteilung, auf Entstehung der verschiedenen Farben und auf die Mendelschen Regeln eingegangen werden. Dazu wird auf die entsprechende Literatur verwiesen.
Trotzdem müssen zuvor auch an dieser Stelle ein paar Begriffe und Regeln festgelegt werden.
Schreibweisen der Farbnamen und Erbformeln
Leider finden wir auch in deutschsprachiger Literatur die englische Schreibweise von Farbnamen und Farbeigenschaften, die nur kleingeschriebene Wörter enthalten. Das widerspricht den Regeln unserer deutschen Orthografie! Eine Farbe wird großgeschrieben (das Rot), ein Halbedelstein wird großgeschrieben (der Achat) weil es Substantive sind. Ein Adjektiv hingegen wird kleingeschrieben (intensiv, dominant).
Weiter ist festgelegt:
- Das Genortsymbol bekommt Buchstaben (maximal drei) die vom englischen Namen der Mutation abgeleitet ist.
- Vererbt die Mutation dominant, wird der erste Buchstabe großgeschrieben. Rezessive Mutationen bekommen kleine Buchstaben als Symbol. Das gilt dann auch für das entsprechende unmutierte Wildgen!
- Wenn ein Gen mehrfach mutiert, werden die mutierten Allele mit kleinen Buchstaben gekennzeichnet und unmittelbar hinter das Genortsymbol hochgestellt angefügt (oox). Wenn hinter dem Genortsymbol ein hochgestelltes “+” geschrieben wird, steht das für ein unmutiertes Gen (o+). Unmutierte Gene werden textlich auch mit „nicht“ oder „non“ bezeichnet (nicht Opal, non-opal).
- Symbole gekoppelte Gene – Gene befinden sich auf dem gleichen Chromosom – werden mit einem “_” (Unterstrich) verbunden (s+_dr).
- Genotypen werden stets in Schrägschrift (kursiv) geschrieben.
Siehe dazu auch Dirk Van den Abeele: „Internationale Namensgebung“ der MUTAVI Research & Advice Group.
Für den „Hausgebrauch“ ist es jedoch vollkommen unerheblich, welche Buchstaben verwendet werden. Wichtig ist, dass man die Vererbungsweise einer zu betrachteten Eigenschaft kennt.
Vererbungsmechanismen
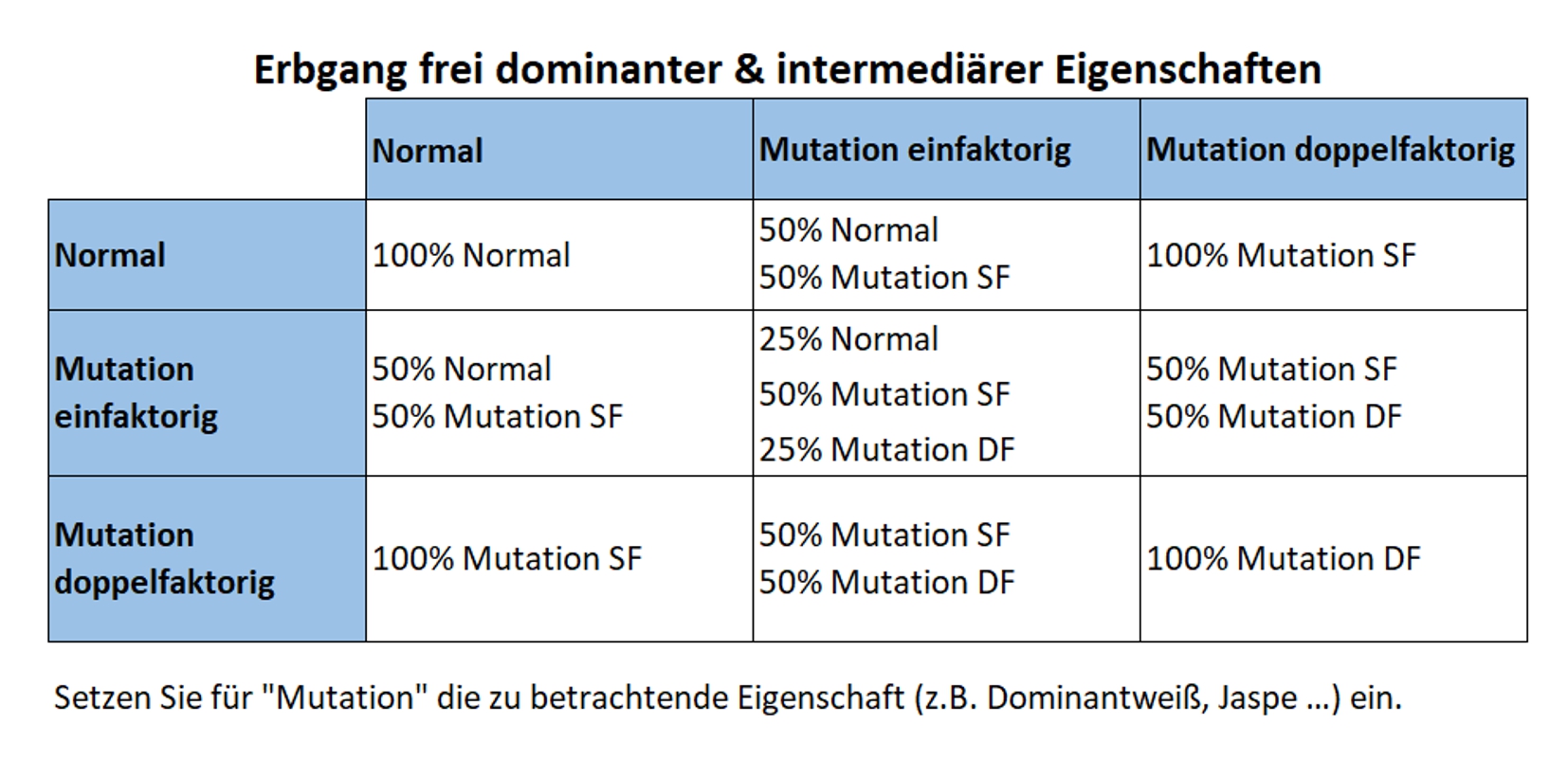
1. Freie rezessive Vererbung
Die betrachteten Gene liegen nicht auf einem Geschlechtschromosom, sondern auf einem der anderen („freien“) Chromosome, die auch Autosome genannt werden. Diese Vererbunsweise wird im Züchterjargon meist nur als „rezessiv“ bezeichnet. In der englischen Sprache ist es „non-sex-linked recessive“ oder „NSL rezessive“.
Eine rezessive Eigenschaft kann nur dann im Phänotyp sichtbar werden, wenn beide Allele eines Gens die Mutation besitzen. Trägt nur ein Allel die Mutation, sprechen wir von spalterbigen Vögeln (Normal / Mutation). Bei manchen Farbeigenschaften kann die Spalterbigkeit von versierten Züchtern optisch erkannt werden, da es feine Farbunterschiede geben kann. Das ist ein Hinweis darauf, dass die Eigenschaft unvollständig rezessiv vererbt.

2. Freie dominante Vererbung
Die betrachteten Gene liegen nicht auf einem Geschlechtschromosom, sondern auf einem der anderen („freien“) Chromosome, die auch Autosome genannt werden. Sie wird im Züchterjargon meist nur als „dominant“ bezeichnet. In der englischen Sprache ist es „non-sex-linked dominant“ oder „NSL dominant“.
Wenn nur ein Allel des Gens die Mutation trägt, sprechen wir von einfaktorigen Vögeln (SF – single-factor). Diese zeigen bereits die Mutation im Phänotyp. Wenn beide Allele des Gens die Mutation tragen, sprechen wir von doppelfaktorigen Vögeln (DF – double-factor). Diese zeigen die Mutation deutlich ausgeprägter (siehe u. a. Jaspe-Kanarien).
Bei manchen dominant vererbenden Eigenschaften (z. B. Dominantpastell und Wange beim Zebrafink, Federhaube bei Zebrafink, Mövchen und Kanarien, Dominantweiß bei Kanarien) sind doppelfaktorige Vögel nicht oder nur eingeschränkt lebensfähig. Sie sterben bereits im Ei oder kurz nach dem Schlupf. Grund ist ein Letalfaktor, der nur bei Homozygotie wirksam wird.
Zur Vermeidung homozygoter Individuen dürfen niemals zwei Merkmalsträger miteinander verpaart werden!
Einfaktorige Vögel sind jedoch ohne Einschränkung vital und fruchtbar.
2a. Freie intermediäre Vererbung
Beim intermediären Erbgang liegt der Phänotyp einfaktoriger Nachkommen zwischen „Normal“ und den doppelfaktorigen Vögeln. Die Dominanz der Mutation ist also nicht vollständig, weshalb man auch von einer „unvollständigen Dominanz“ spricht.
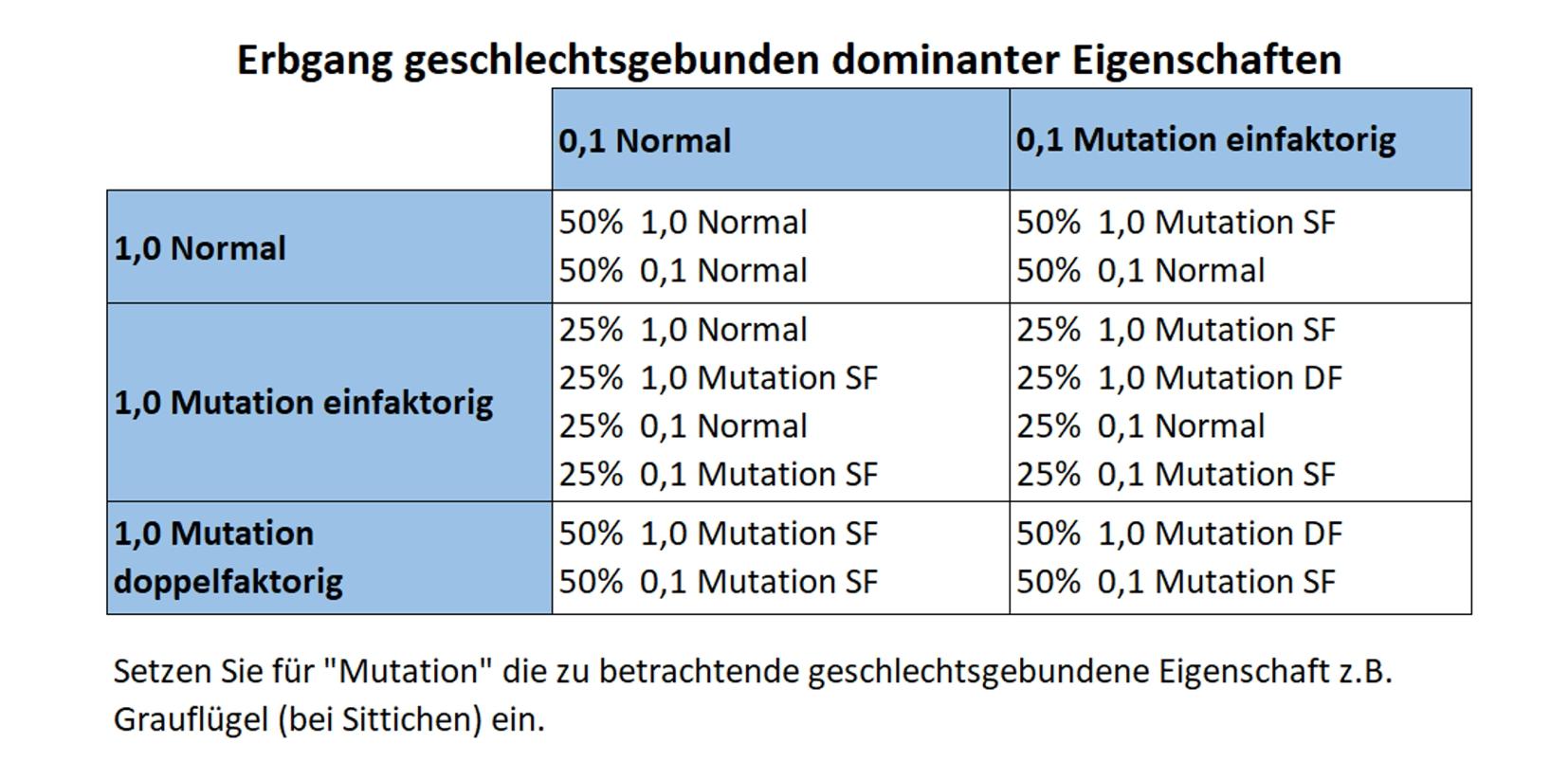
3. Geschlechtsgebunden rezessive Vererbung
Die betrachteten Gene liegen ausschließlich auf dem Geschlechtschromosom, dem Z-Chromosom. Diese rezessive Vererbungsweise ist bei Kanarien bisher die einzige geschlechtsgebundene Vererbung und wird deshalb von Kanarienzüchtern auch meist nur als „geschlechtsgebunden“ bezeichnet. In der englischen Sprache ist es „sex-linked recessiv“ oder „SL recessiv“.
Eine geschlechtsgebunden-rezessive Eigenschaft kann nur dann im Phänotyp der Männchen sichtbar werden, wenn beide Allele des Gens die Mutation besitzen. Ist nur ein Allel des Gens mutiert, sprechen wir von spalterbigen Männchen (Normal / Mutation). Bei manchen Farbeigenschaften kann die Spalterbigkeit von versierten Züchtern optisch erkannt werden, da es feine Farbunterschiede geben kann. Das ist ein Hinweis darauf, dass die Eigenschaft unvollständig rezessiv vererbt.
Da Vogelweibchen nur ein Z-Chromosom besitzen, können sie niemals spalterbig in einer geschlechtsgebundenen Eigenschaft sein. Allerdings reicht dieses eine mutierte Gen aus, um bei Weibchen die geschlechtsgebundene Eigenschaft im Phänotyp auszubilden.

4. Geschlechtsgebunden dominante Vererbung
Diese ist bei Kanarien bisher noch nicht aufgetreten, soll aber hier der Vollständigkeit wegen aufgeführt werden, da sie u. a. bei Sittichen auftritt (z. B. dominant Grauflügel). In der englischen Sprache ist es „sex-linked dominant“ oder „SL dominant“.
Bei einer geschlechtsgebunden dominanten Vererbung kann es doppelfaktorige (DF – double-factor) und einfaktorige (SF – single-factor) Männchen geben.
Weibchen sind in diesem Fall immer nur einfaktorig.
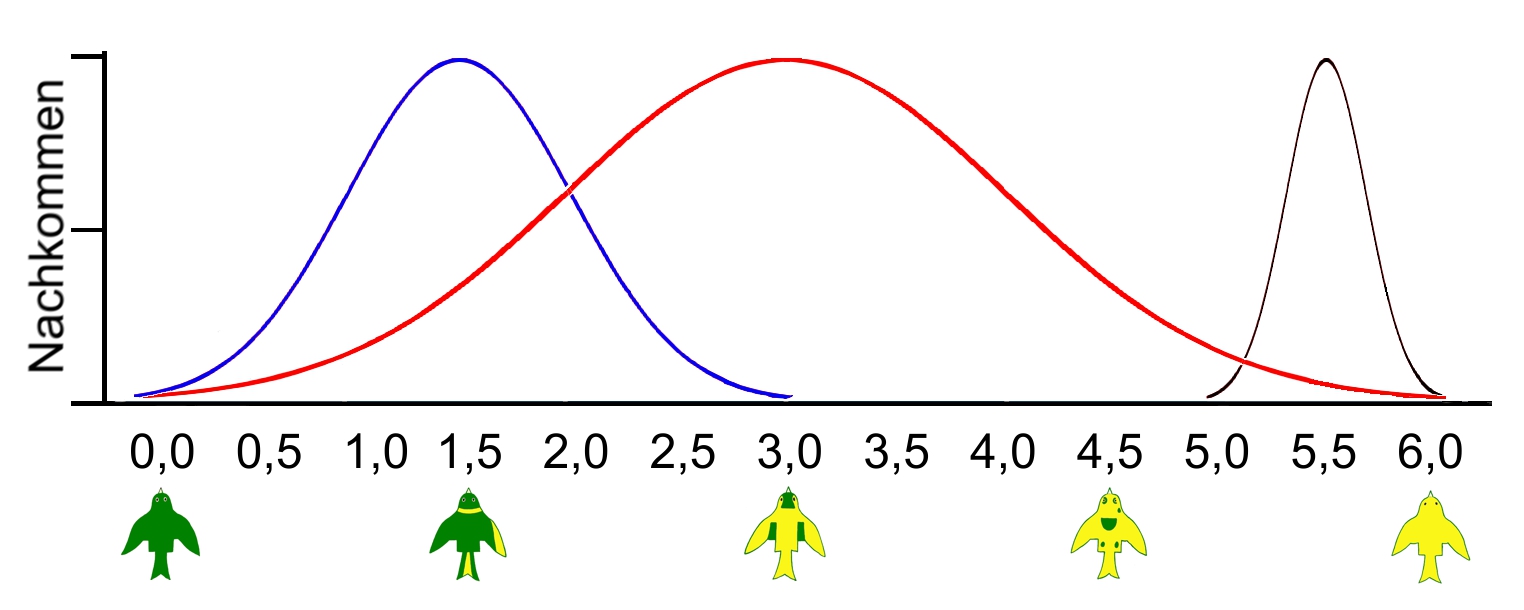
Quantitative Vererbung
Die quantitative Genetik befasst sich mit den erblichen Komponenten von Merkmalen, die durch das Zusammenwirken vieler Gene bestimmt werden, und die auf einer Skala gemessen werden können. Quantitative Merkmale sind all jene Merkmale, die durch fortwährende Selektion kontinuierlich in die eine oder andere Richtung beeinflusst werden können. Das sind z. B. die Körpergröße, das Körpergewicht, die Körperform, die Federlänge, die Federtextur, die Anzahl gelegter Eier oder der Scheckungsgrad. Zusätzlich können diese Merkmale durch äußere Umwelt- und Ernährungsbedingungen beeinflussbar sein
Die Varianz der zu betrachtenden quantitativen Merkmale kann man statistisch berechnen und in einer Kurve darstellen. Anhand dieser Normalverteilungskurve ist die Wahrscheinlichkeit der zu erwartenden Merkmale der Nachkommen ablesbar.
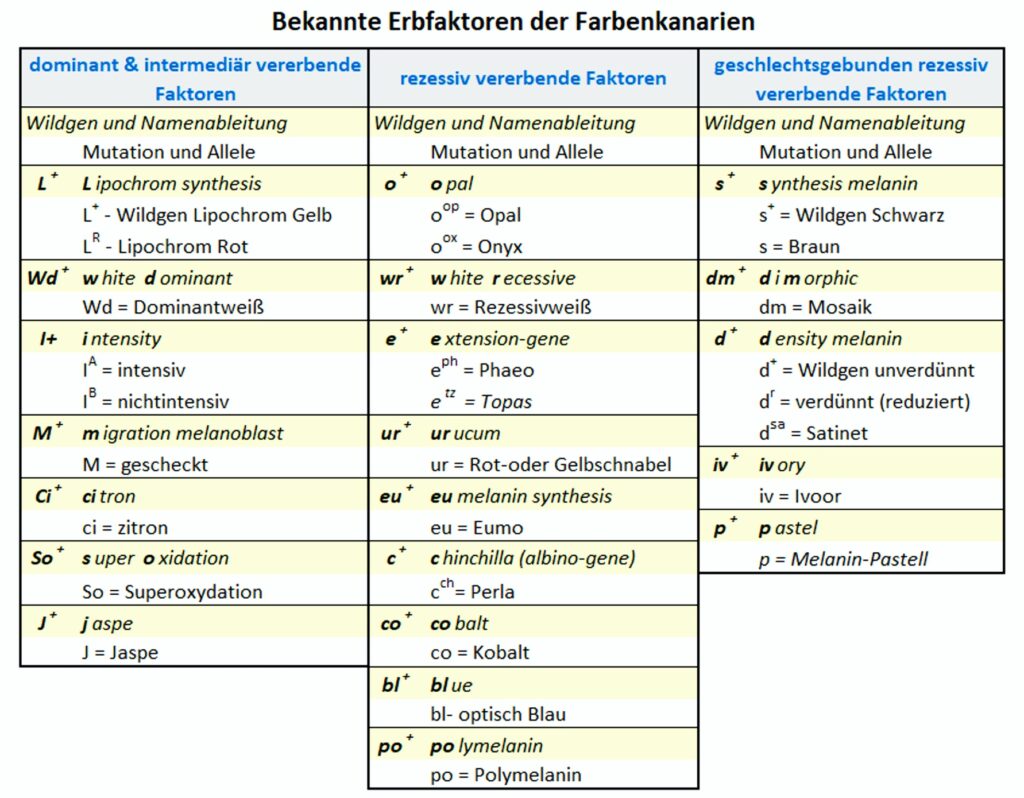
Übersicht bekannter Erbfaktoren
Quellen und Literatur
Arbeitsgruppe AZ-AGZ Farben/Genetik. Unter: https://az-agz-farben-genetik.jimdofree.com
N. Schramm: Kompendium-Kanarien, Band 2, Genetik, Gefiederfärbung, Farbenkanarien, Mischlinge. Books on Demand, Norderstedt, 2018. Siehe auch hier